
抗体的结构和片段
信息来源:金开瑞 作者:genecreate 发布时间:2019-01-15 15:28:19
常规抗体
常规的抗体或者是完整大小的抗体是被称为免疫球蛋白的糖蛋白,是浆细胞对外来分子或者抗原反应后产生的。抗体最基础的功能是结合特异性抗原并激发免疫反应,从而保护机体免于感染。抗体包括好几种亚型,这里主要是介绍IgG和IgM亚型的抗体。IgG和IgM亚型的抗体被广泛应用于科研、诊断和治疗等方面。
1.常规抗体的结构
一个完整的抗体的基础单元结构有四条肽链组成,包括两条重链和两条轻链,并通过二硫键结合在一起。抗体的形状像一个Y字母,Y结构的铰链区具有弹性。每条肽链就有一个恒定区(在所有的抗体中都非常的保守)和一条可变区(在一个种抗体中都是特异性的)。轻链可变区的标示符号是VL,而轻链恒定区的标示符号是CL(图一左)。 类似的,重链的可变区和恒定区分别被标示为(VH)和(CH)。碳水化合物通常和重链的CH2区域结合。Fc段只包括重链的恒定区(CH), 但是和抗原结合的Fab段(Fab)包括一个恒定区和重链的可变区以及轻链的可变区(VH and VL)。 Fv区域(variable fragment) 只包含两个可变区
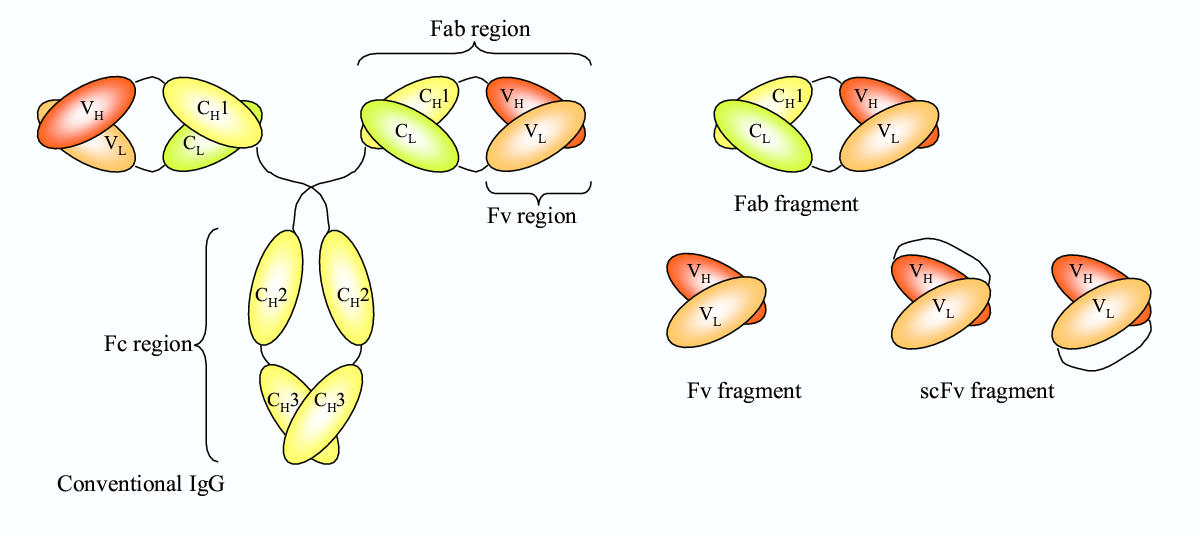
一个完整的常规抗体(左)和通常的抗体片段(右)的基本结构
2.常规抗体的应用
常规抗体在科研中用于通过蛋白质印迹、免疫组织化学以及酶联免疫吸附法等方法检测目的蛋白已经有几十年了。完整大小的抗体还被应用于临床的检测,比如妊娠试验以及用ELISA检测血液中HIV病毒。另外,常规的完整的抗体还被应用于疾病的治疗。例如,英利昔单抗是一个可以识别肿瘤坏死因子的抗体,被用于治疗肠道疾病和类风湿性关节炎。 曲妥珠单抗或者赫赛汀是一种可以结合上皮生长因子二的抗体,被用于治疗转移的乳腺癌。 另外,还有好多抗体,包括Muromomab,被用于器官移植后的基础治疗来防止发生移植物排斥反应。
用常规抗体的优点包括Fc区域能够激活机体的免疫反应并结合到目标分子上使之被破坏。用完整抗体的缺点包括由于其分子太大不能被渗入到某些组织中。 Fc段能够激活一些有害于病人的免疫反应是临床上应用完整抗体治疗的一个缺点。Fc区域通常能引起一些非特异性的结合,对抗体的检测的应用不利。
抗体片段
一个抗体的片段可以通过化学试剂和基因工程的方法获得。化学试剂片段通过是通过打断铰链区的二硫键或者是用蛋白酶,包括胃蛋白酶和木瓜蛋白酶,消化抗体后产生的。基因工程产生的片段提供大量的片段,每一个都能有特异性的结合区和功能特质。
Fab, Fab', (Fab')2, 和 Fv
抗体经过化学试剂处理的和蛋白酶消化可以得到抗原结合片段(Fab), 其来源于IgG 和 IgM 亚型抗体的可变区。 去掉Fabs片段的抗体部分就是Fc片段了,由重链的恒定区组成。 抗原结合片段包括了Fab, Fab', (Fab')2, 和 Fv。 这些片段能够结合抗原,但是它们缺少Fc段,其包括重链的恒定区2和3。当抗体被木瓜蛋白酶消化后,两个独立的F(ab)片段从Fc区域分离下来。然而用胃蛋白酶消化后,一个带有小部分的Fc铰链区的 F(ab')2 片段从抗体中分离下来。尽管利用化学试剂的方法分离抗体的片段能够产生很多有用的诊断和治疗的工具,但是是非常费时的而且需要大量的抗体作为原材料。
单价的F(ab) 片段只有一个抗原结合区,然而多价的F(ab')2 片段有两个抗原结合区,它们通过二硫键结合在一起。F(ab')2 片段产生2个单价的Fab' 片段和一个游离的硫基,其能够用于其他分子的结合。
Fv 片段是IgG和IgM类型抗体通过酶法分析后产物中最小的片段。Fv片段抗原结合区,其由VH和VC区域组成,但是他们缺少CH1和CL区域(Figure 1 right panel)。VH 和VL 通过非共价键在Fv 片段里结合在一起。
ScFv, 双特异抗体,三特异抗体,四特异抗体,双-scFv,mimi抗体,Fab2,Fab3
基因工程方法能够产生单链可变区(ScFv),它是Fv类型片段,其包括了通过易弯曲多肽链接在一起的VH和VL区域(图一右)。如果结合区至少有12个残基的长度,ScFv片段就是单抗。 通过操作V-结构域和铰链区的长度可以创作不同形式的Fv分子。 连接体是3-11残基的产生的scFv分子不能折叠成有功能的Fv结构域。这些分子和其他的scFv分子一起可以创作出一个双价的双特异性抗体。 如果连接体的长度小于3个残基,scFv 分子相互作用后能够产生三特异性或者四特异性抗体。多价的scFvs与对应的单价的抗体相比具有更强的结合抗原的亲和力。 Mini抗体是scFv-CH3融合蛋白,被装进到双价的二聚体中。 Bis-scFv 片段是双特异性的。 小型化的ScFv片段能够通过两个不同的可变区产生,能够让这些Bis-scFv分子同时结合两个不能的表位。基因学方法能够用来产生双特异性的Fab 二聚体(Fab2)和三特异性的Fab 三聚体(Fab3)。这些抗体片段同时能够结合2 (Fab2)或者3(Fab3)不同的抗体。
骆驼/鲨鱼源抗体和纳米抗体
除常规抗体外,骆驼科动物和鲨鱼还含有一些特殊的重链抗体(hcAb),它们完全由缺乏轻链的重链同源二聚体组成。这些抗体的Fab部分称为VHH(重链抗体的可变结构域),是天然发现的最小的抗原结合区域。纳米抗体是VHH衍生的重组结构域且能结合抗原。它们非常稳定,可以很容易地通过传统的简单系统大量生产,如细菌(但具有轻链和重链的常规抗体难以在细菌系统中表达),因此是一种对研究和治疗应用有前景的工具,特别是在超分辨显微镜,质谱和靶向蛋白质降解领域。纳米抗体可以通过与多肽共价连接被递送到活细胞内,或者直接在体内表达并识别靶点,但同时具有轻链和重链的常规抗体不能用于活细胞。例如,抗GFP纳米抗体用于开发研究体内神经元活性的电磁控制系统。抗RFP或GFP的纳米抗体与远红染料Atto结合时,可以获得超过GFP或RFP 118倍的荧光信号放大,可用于产生全身小鼠神经元连接。它们还可在结构研究中用来稳定蛋白质的活性状态。现已证明,AAV表达载体能生产通用流感疫苗,这个载体有抗四种不同流感病毒株的连锁纳米抗体。重组抗小鼠和抗兔IgG二级纳米抗体有可能取代广泛使用的多克隆二抗。纳米抗体具有跨越血脑屏障的独特能力; 然而,往往会很快被处理和清除出体外。纳米抗体可用于一些特定目的,如(共)免疫沉淀或活细胞中细胞内靶标的实时荧光蛋白示踪。
牛超长CDR3H
大约10%的牛免疫球蛋白包含一个具有多个半胱氨酸残基的长CDR3H区域,这种长茎结构被认为有助于抗体多样性。
抗体片段的应用
在一些应用中,片段比完整的抗体更加有优越性。这个话题最近被Nelson写了综述。 其中一个优越性是片段比完整的抗体要小,能够进入完整抗体进不去的组织中发挥治疗作用和进行免疫组织化学染色。 抗体片段比常规的抗体更小,通常没有被糖基化,可以使它们的产物在原核表达系统中表达,从而节省时间和资金消耗。然而, 缺少Fc结构域的片段在体内比常规抗体降解的速度要快,而且不能够激发Fc介导的细胞毒性进程,除非它们结合一个为了达到更好治疗目的的有效的原件。然而,缺少Fc结构域对于免疫组织化学和其他的检测又是有利的,因为减少了抗体与Fc受体的非特异性结合。没有Fc区域的抗体片段具有能够减少非特异性结合的优点。抗流感的神经氨酸酶抗体 NC10 是一个在诊断中被广泛应用的一个ScFv。抗上皮细胞粘附分子Ep-CAM抗体MOC-31 是一个用于癌症治疗的ScFv。 双特异性抗体,三特异性抗体和四特异性抗体在放射免疫治疗和体内的影像学诊断方面都有潜在的应用价值。
尽管各种抗体片段具有某些优点,但它们通常不用于实验。在截止2016年3月来邦网调研的38430篇文献中,只有少数文章与抗体Fab片段的应用有关。Roche抗地高辛配基Fab片段在原位杂交实验中以1:1000或1:2000稀释使用,结果表明主要的近端肢体细分取决于可扩散信号,纤毛发生由进化组装的顺式调节元件调控,蝴蝶中的模拟翼模式进化可由optix基因调控。他们还用来进行蛋白质折叠分析,通过单分子力谱以研究单个钙调蛋白分子折叠的过程。 Roche抗地高辛配基抗体在绵羊中产生并通过木瓜蛋白酶消化生产。Roche抗荧光素碱性磷酸酶Fab片段用来进行Southern印迹以研究拓扑异构酶II的功能。Roche抗地高辛配基或抗荧光素Fab试剂的最新研究实例包括,描述神经元表达的连接蛋白35b的表达模式,讨论果蝇胚胎,卵巢和培养细胞系中凋亡性细胞死亡的方法,测试突触GluA2浓度是否能够调节蛋白质运输或局部翻译,研究人类皮质形成与受损的音猬信号传导之间的关系,研究发育过程中时钟基因表达行波的机制,研究TRF2与核纤层蛋白A/C之间的相互作用如何调控染色体结构,描述一种连接小鼠神经元特定亚型的发育途径的检查方法及其它应用。
来自Invitrogen公司的Alexa 546共价结合的驴抗兔IgG Fab片段,用在免疫组化实验中来研究sFLT-1在维持小鼠模型中无血管感光细胞层中的作用。该公司的Alexa Fluor 488兔抗山羊IgG(H+L)(Fab')2片段,被用在免疫组化实验中来研究听觉毛细胞中尖端连接再生的机制。
Fc片段受体
Fc受体(FcR)是主要在先天免疫细胞上/中表达的分子,能识别并结合抗体的Fc结构域,从而为它们提供细胞系统以引发免疫应答。FcR的多种功能反映了抗体广泛的保护或调控作用,包括介导靶向底物的中和和清除以及适应性免疫的编程。FcR的生物学功能受基于免疫受体酪氨酸的活化基序(ITAM)和基于免疫受体酪氨酸的抑制基序(ITIM)的调控,分别作为激活和抑制信号传导途径的受体界面。因此,ITAMs的信号传导可以引发细胞活化,吞噬和内吞作用,而ITIMs的信号传导对细胞活化具有抑制作用。现已有关于所有类别的免疫球蛋白的FcR的描述,下面将对其中一些进行讨论。
IgG受体
该家族包括FcγRI,FcγRII,FcγRIII及其亚型。它们负责抗体依赖性细胞介导的细胞毒性(ADCC)和抗体依赖性细胞介导的吞噬作用(ACDP)。
另一种结合IgG的受体是新生儿Fc受体(FcRn),其参与从母体到胎儿的被动体液免疫的转移。FcRn还可以保护IgG免于体内降解,这是它们在血清中的半衰期长的原因。通过Fc区的改变以促进Fc-FcRn相互作用,该现象产生了更好的治疗性抗体的开发。
IgE受体
它们包括能够结合单体IgE的高亲和力FcεRI和能够优先与IgE复合物相互作用的低亲和力C型凝集素FcεRII。FcεRI通过刺激细胞脱粒和在肥大细胞和嗜碱性粒细胞上释放一系列炎症介质来介导许多过敏反应的即时超敏应答。FcεRII既能以膜结合形式存在用于递送下调IgE合成的信号,又能以可溶性片段形式存在产生上调效应。它在人类气道和肠上皮细胞中IgE过敏原复合物胞吞过程中的作用正在被积极地研究,可能是治疗食物过敏引起的过敏性气道炎症的潜在靶点。
IgA受体
FcαRI是该类型的唯一成员,仅在骨髓谱系细胞中表达。根据IgA结合状态,它在促炎症和抗炎症反应中起作用。虽然结合于粘膜部位的分泌型IgA(SIgA)具有抗炎作用,包括预防病原体入侵,血清IgA的结合导致炎症反应。 FcαRI还根据炎症微环境调节中性粒细胞活力。
TRIM21
TRIM21显示出非常广泛的抗体特异性,因而可以与其他FcR区分开。它可以结合IgG,IgM和IgA。此外,它在大多数产生组织的细胞中表达。TRIM2通过靶向细胞溶质病毒-抗体复合物进行蛋白酶体降解,参与抗体介导的病毒复制干扰。
Fc结构域与FcR的结合可能在基于单克隆抗体的分析方法中具有负面效果,例如免疫组织化学(IHC),流式细胞术(FACS)和染色质免疫沉淀(ChIP)。与FcR的非特异性结合可能引入背景噪声,从而导致假阳性。该问题的解决方案包括使用(i)用于门控的同种型对照,(ii)血清来广泛竞争参与非特异性结合的受体,或(iii)纯化的IgG以特异性阻断Fc受体。Innovex 的Fc受体阻断剂#NB309可在IHC或IC实验中用于阻断石蜡或冷冻切片BD Biosciences#553142可用于流式细胞术。
上一条:柱层析蛋白质纯化方法学介绍
下一条:microRNA实验方法
最新动态
-
04.22
一文读懂EMSA技术核心要点,让“emsa” 秒变“easy”
-
04.02
4·2世界孤独症日 | 聆听“星”声“泌”语,让爱来,让碍走
-
04.01
酵母杂交核心技术:深度Q&A帮你轻松突破实验瓶颈
-
03.19
【客户文献解读,IF>11】食管癌的"隐形推手":MALR-ILF3-HIF1a轴的强大作用
-
03.18
siRNA介绍及药物研发的现状前景
-
03.18
知无不“研”|5篇高分文献带你一览高通量酵母杂交的非凡魅力~
-
02.27
【客户文章分享】SHMT2 通过 5′UTR 依赖性 ADAM10 翻译启动介导小分子诱导的阿尔茨海默病病理学缓解过程
-
01.24
客户文献解读 | 中医为什么能治流感?是玄学还是运气?INT J NANOMED揭示鱼腥草抗病毒机制及范围!
-
01.24
客户文献分享,IF>11|Shank3:脑缺血再灌注损伤的守护者,揭示神经保护的新篇章
-
12.27
文献解读 | 高密度脂蛋白通过miR-181a-5p调控自噬影响血管新生
 X
X 






