
【客户文献解读,IF>11】食管癌的"隐形推手":MALR-ILF3-HIF1a轴的强大作用
肿瘤相关巨噬细胞(TAM)是肿瘤微环境中数量最多的免疫细胞,对肿瘤的发生发展起到重要调节作用。作者发现了一种与巨噬细胞相关的长非编码RNA(lncRNA),名为MALR,它可以促进食管鳞状细胞癌(ESCC)的进展。TAM通过分泌TNFa上调ESCC细胞中MALR的表达,MALR可以通过激活HIF1a信号通路促进糖酵解和血管生成。在机制上,MALR与ILF3的dsRBD1结构域结合,促进ILF3蛋白的稳定性以及由ILF3介导的液-液相分离(LLPS),从而防止PARN介导的降解,提高HIF1a mRNA的稳定性。如果MALR缺失,基于细胞系的以及患者来源的异种移植肿瘤的生长会被抑制。在临床上,MALR的高表达与HIF1a靶基因的表达正相关,并预示着食管癌患者的预后不良。总的来说,这项研究揭示了MALR/ILF3介导的LLPS在肿瘤微环境重塑中的生理作用,突出了MALR-ILF3-HIF1a轴作为癌症治疗的潜在靶点。
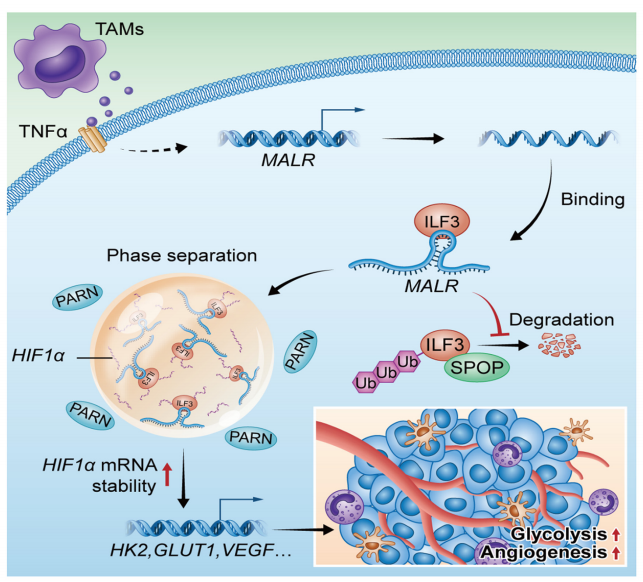
01 TAMs通过TNFa信号通路上调MALR的表达
作者首先从与TAMs共培养的ESCC细胞中分离RNA,并进行RNA-seq分析以确定TAM上调的lncRNA。通过比较与TAMs共培养的ESCC细胞和单独培养的ESCC细胞,确定了前20个TAM上调的lncRNA(图1A)。为了进一步缩小候选者范围,作者对ESCC肿瘤组织与其配对的正常组织(N=20)进行了RNA-seq分析,并确定了前20个在肿瘤组织中上调的lncRNA(图1B)。在上调的lncRNA中,只有一个候选者在两组中均出现,这个lncRNA被命名为MALR。
通过qRT-PCR分析,证实了MALR在ESCC组织(图1C)以及与来自患者的TAMs、M2型巨噬细胞或THP-1细胞共培养的ESCC细胞(KYSE150和TE-11)中上调(图1D-F)。相关性分析显示,ESCC肿瘤组织中MALR的表达与CD68的表达呈正相关,CD68被认为是巨噬细胞的一种特异性标记(图1G)。此外,ESCC患者中MALR高表达者总体生存率和无病生存率较差(图1H和I),这表明MALR是ESCC潜在的预后生物标志物。此外,当与TNFa敲低的TAMs共培养或用中和抗体阻断TNFa时,肿瘤细胞中MALR的上调没有发生显著变化(图1K;补充图S1I),这表明TNFa是导致TAMs诱导MALR表达的主要炎症细胞因子。综上,这些结果表明TAMs通过TNFa信号通路上调MALR的表达。
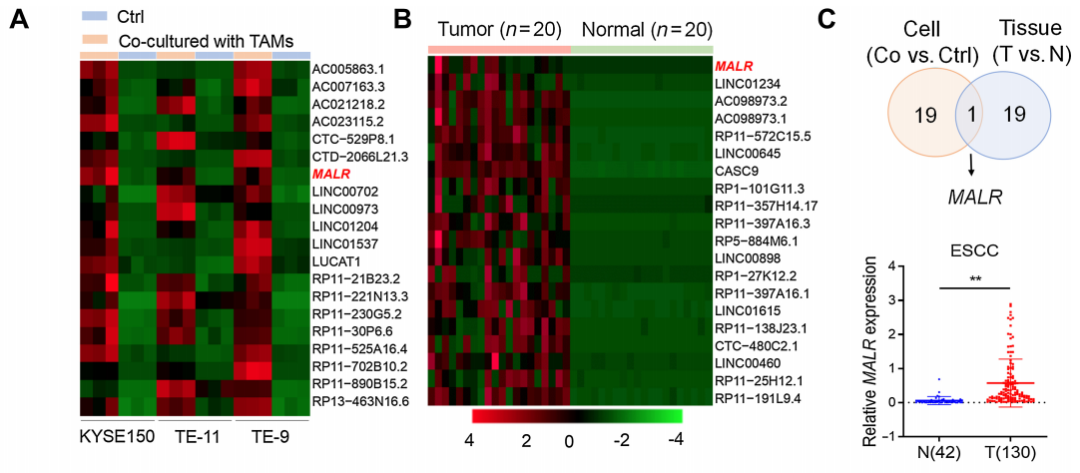
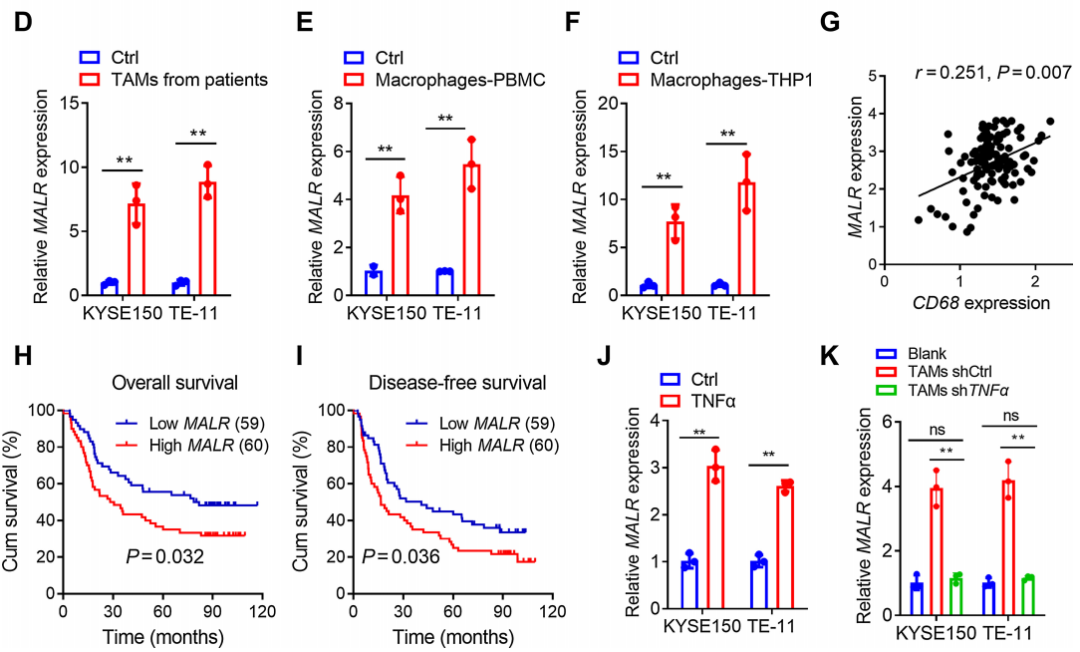
图1. MALR通过TAMs分泌的TNFa上调
02 MALR通过激活HIF1a通路促进ESCC细胞的恶性表型
考虑到MALR可能参与ESCC的恶性发展,作者在两个高表达MALR的细胞系中(TE-15和TE-9)构建了MALR敲除(KO)的ESCC细胞,并进行了RNA-seq分析以确定受MALR调节的靶基因。热图结果表明,在两种ESCC细胞系中,当MALR敲除后HIF1a的表达显著下调(图2A)。据报道,HIF1a在缺氧条件下调控许多基因以刺激糖酵解代谢和血管生成。对TCGA数据的基因集富集分析表明,MALR高表达与缺氧、糖酵解和血管生成呈正相关(图2B)。qRT-PCR分析显示,在缺氧条件下培养的MALR敲除细胞中,HIF1a靶基因的转录显著减少(图2C),这一点也得到了免疫印迹(IB)分析的证实(图2D)。这些数据表明MALR在癌细胞中可能激活HIF1a信号通路。
MALR的缺失显著削弱了ESCC细胞在缺氧条件下培养时的糖酵解活性和细胞生长(图2E和F;补充图S2B)。VEGF是癌症中关键的促血管生成因子,对癌症进展至关重要。正如ELISA所证实的那样,在缺氧条件下培养24小时后,MALR的缺失显著降低了培养基中VEGF的分泌(图2G)。此外,当人脐静脉内皮细胞(HUVEC)在从MALR敲除细胞获得的培养基中培养时,管状形成能力显著受损(图2H)。TNFa刺激显著增强了ESCC细胞在缺氧条件下培养时的糖酵解活性、细胞生长和管状形成能力,而这种作用可被MALR敲除所减弱(图2I-K)。综上所述,这些数据表明MALR表达与HIF1a激活之间存在强相关性,并强调了MALR在重编程癌细胞代谢和促进血管生成中的潜在作用。
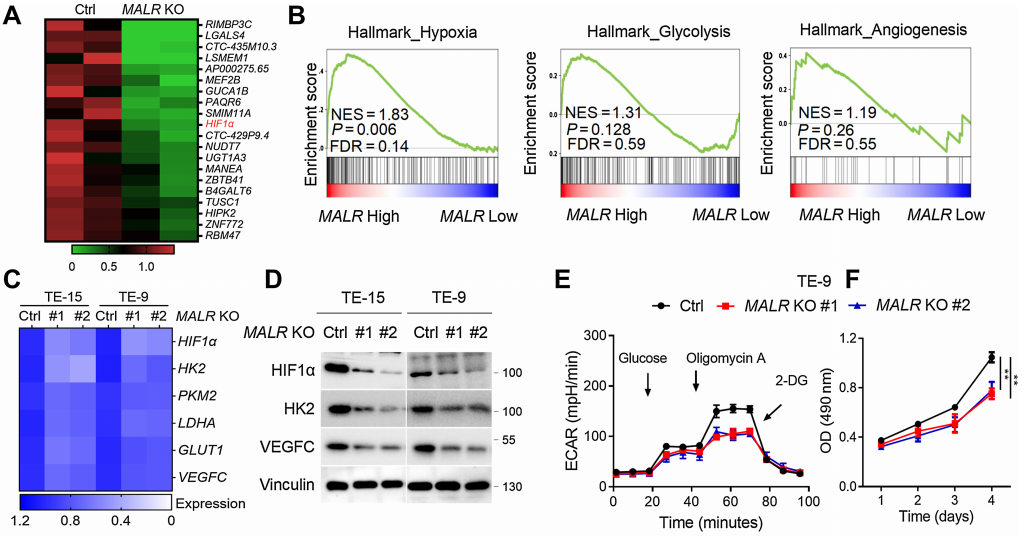
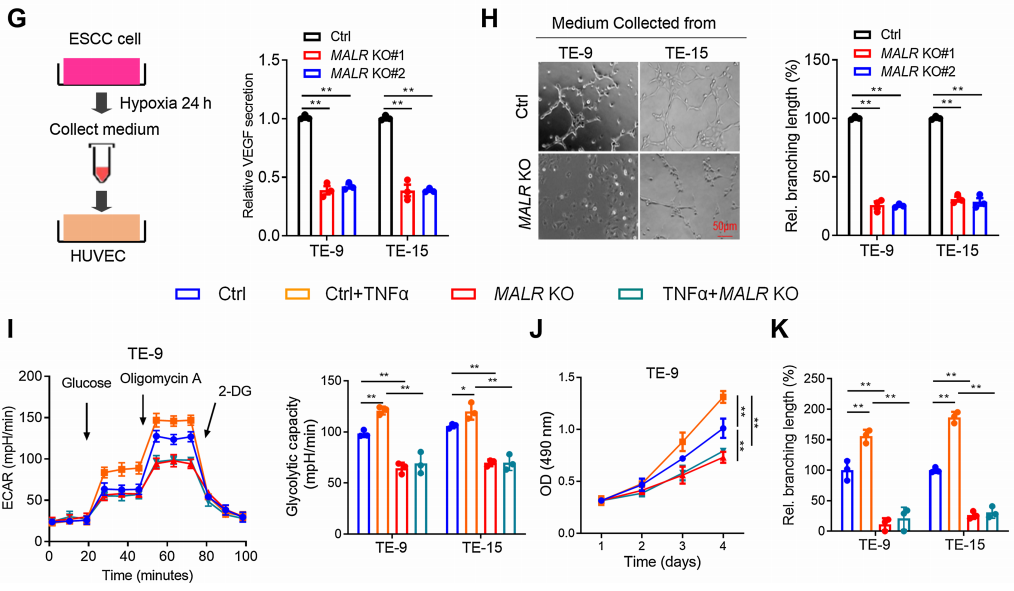
图2. MALR通过激活HIF1a通路促进ESCC细胞的恶性表型
03 ILF3与MALR结合并经历LLPS
接下来,作者通过FISH检测和MALR的亚细胞定位发现MALR主要定位于细胞核(图3A和B;补充图S3A)。为了探究MALR在肿瘤进展中的潜在作用和机制,作者进行了RNA pull-down分析,随后通过MS鉴定了与MALR相关的蛋白质,如之前报道的那样,特异性与ILF3结合是正义MALR,而不是反义MALR(图3C)。MALR与ILF3之间的特异性相互作用也通过RIP实验得到证实(图3D;补充图S3B)。进一步的MTRAP实验表明,与阴性对照相比,MS2-MALR和MCP-3FLAG质粒的共表达导致ILF3的明显富集(图3E)。作者还通过ChIRP分析在体内验证了MALR与ILF3的相互作用(图3F)。
此外,作者进行了CLIP-PCR分析,并鉴定了MALR的T3片段介导了MALR与ILF3的相互作用(图3G)。通过进行ILF3蛋白序列的生物信息学分析,作者发现ILF3的C端高度无序,并包含IDRs(图3H)。研究表明IDRs经常出现在相分离的隔室中,因此作者推测ILF3分子有可能发生LLPS。为了评估与MALR的相互作用是否诱导ILF3发生LLPS,作者将EGFP-ILF3蛋白(绿色)与Alexa Fluor 546-14-UTP标记的MALR(红色)混合。混合后,ILF3和MALR形成了微米大小的液滴(图3I),这在ESCC细胞中通过FISH和IF检测也观察到了(图3J)。FRAP分析进一步证明了ILF3介导的LLPS的动力学特性(图3K;补充影像S1)。如FRAP分析所示,液滴动态地改变,分子在液滴与周围溶液之间进行交换(图3K;补充影像S1)。综上所述,这些结果表明ILF3蛋白能够发生LLPS。
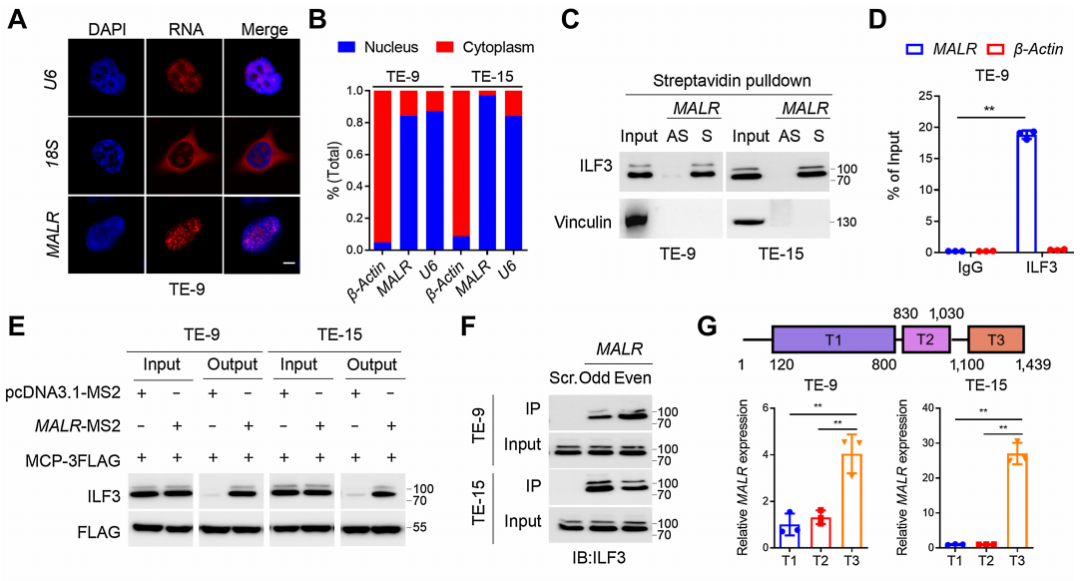
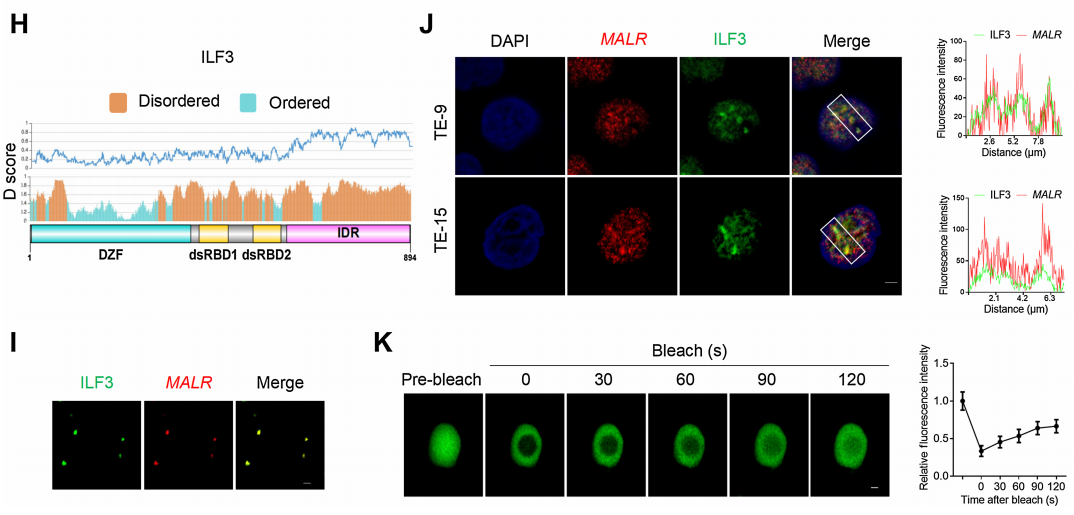
图3. ILF3与MALR结合并经历LLPS
04 MALR促进ILF3蛋白的稳定性和ILF3介导的LLPS
随后,作者评估了MALR-ILF3相互作用对ILF3稳定性的功能效应。作者发现在ESCC细胞与巨噬细胞共培养或用TNFa处理后,ILF3的表达增加(图4A;补充图S4A)。具体来说,MALR KO ESCC细胞中ILF3的表达明显减少,但通过蛋白酶体抑制剂MG-132处理可以恢复(图4B),这表明MALR促进ILF3的稳定性。此外,敲除MALR对ESCC细胞中ILF3的稳定性有显著影响,缩短了ESCC细胞中ILF3的半衰期(图4C和D;补充图S4B)。此外,作者在用FLAG标签标记的ILF3表达的细胞中进行了IP试验,用抗FLAG抗体进行IP检测,并通过IB法检测了泛素水平。结果显示,MALR敲除显著增加了泛素化ILF3的水平(图4E)。这些数据表明,MALR是维持ILF3稳定所必需的。为了找出与MALR相互作用所需的ILF3蛋白的结构域,作者测试了几种截短的ILF3蛋白形式并发现ILF3的dsRBD1片段在TE-9细胞中是必需的(图4F)。综上所述,ILF3的dsRBD1片段是MALR-ILF3相互作用和包括ILF3蛋白稳定性、细胞生长在内的关键功能所必需的。
之前的研究表明,SPOP是一种E3泛素连接酶,通过与dsRBD1片段附近的结构域的物理相互作用来调节ILF3蛋白的稳定性。基于这一发现,作者通过Co-IP实验观察到敲除MALR显著增强了ILF3与SPOP之间的相互作用(图4G),这一结果也得到了Duolink PLA实验的证实(图4H;补充图S4H)。因此,作者的数据表明MALR通过阻止SPOP介导的泛素化来增加ILF3蛋白的稳定性。当ILF3-EGFP蛋白浓度为0.1mmol/L时,在不存在MALR的情况下观察到了液滴的形成(图4I)。此外,正交实验表明ILF3的相分离依赖于MALR的剂量。在存在MALR的情况下,相同蛋白浓度下液滴形成能力显著增强(图4I)。为了进一步探讨MALR对ILF3相分离的影响,作者在ESCC细胞中过表达MALR,发现显著增加了ILF3点状形成(图4J)。MALR KO ESCC细胞中ILF3点状形成受到抑制,这可以通过重新表达FL-MALR来挽救,而在DT3-MALR过表达组中则不是这样(图4K)。这些结果表明,MALR(尤其是T3片段)促进ILF3介导的LLPS形成并促进肿瘤生长。
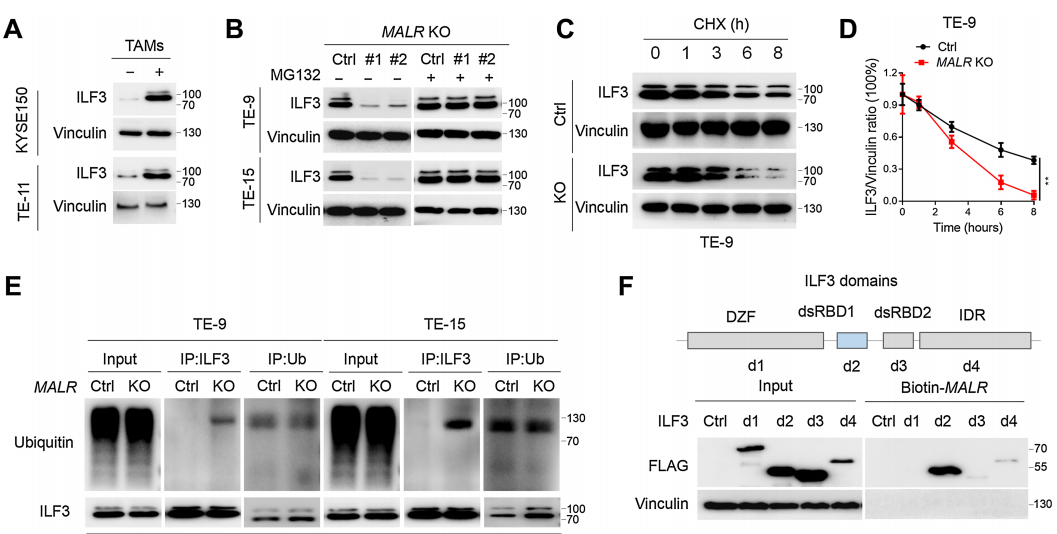
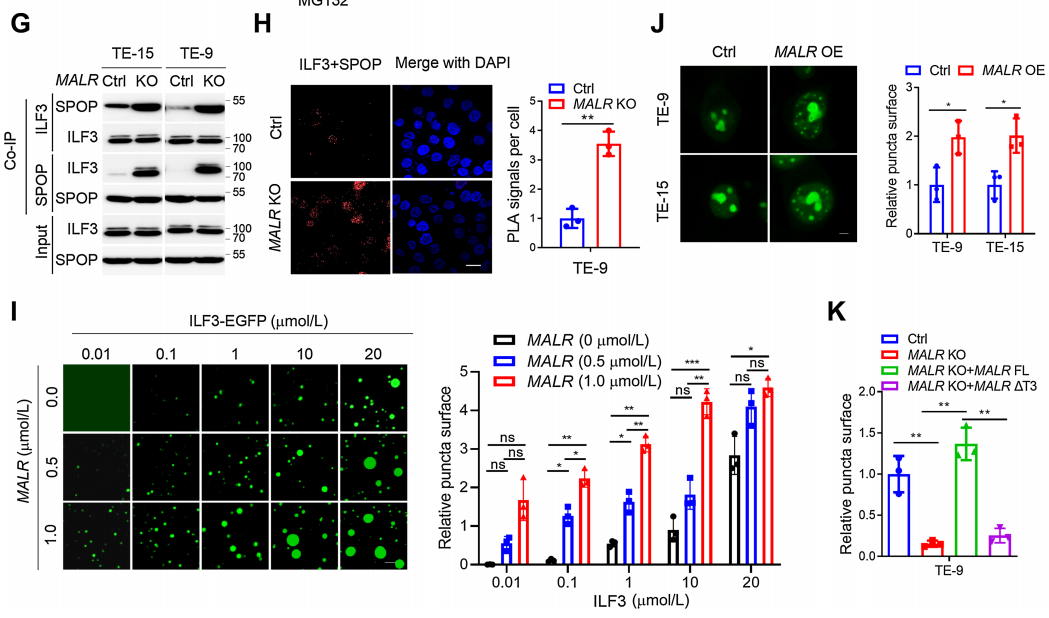
图4. MALR促进ILF3蛋白的稳定性和ILF3介导的LLPS
05 MALR-ILF3介导的LLPS增加了HIF1a mRNA的稳定性
Actinomycin D是一种广泛用于测量mRNA降解率的转录抑制剂,在ESCC细胞中,MALR的缺失降低了HIF1a mRNA的稳定性,而在这些细胞中加入Actinomycin D(图5A;补充图S5B)后也是如此。相反,TNFa处理显著增强了HIF1a mRNA的稳定性,这可以通过在ESCC细胞中敲除MALR来挽救(图5B;补充图S5C)。此外,在MALR KO ESCC细胞中,HIF1a mRNA的稳定性降低可以通过外源性表达WT-ILF3来挽救,但不能通过DIDR-ILF3或DdsRBD1-ILF3来挽救(图5C;补充图S5D)。在MALR KO细胞中,HIF1a mRNA的稳定性降低不能通过与1,6-HD一起使用的WT-ILF3来挽救,却可以通过与对照2,5-HD处理一起使用的WT-ILF3来挽救(图5D;补充图S5E和S5F)。综上所述,这些数据表明MALR-ILF3介导的LLPS在促进HIF1a mRNA稳定性方面起着至关重要的作用。
如图5E所示,作者推断ILF3介导的LLPS可能阻止HIF1a mRNA与PARN之间的相互作用。敲除MALR显著增加了HIF1a mRNA与PARN的相互作用,这通过RIP-PCR测定得到证实(图5F)。作者进一步测试了ILF3 KO ESCC细胞中HIF1a mRNA与PARN的相互作用,RIP-PCR测定表明,敲除ILF3显著增强了HIF1a mRNA与PARN的相互作用,这可以通过在ESCC细胞中过表达shRNA抗性的WT-ILF3来挽救,但不能通过过表达DIDR-ILF3或用1,6-HD处理来挽救(图5G)。总的来说,这些数据表明,MALR-ILF3介导的LLPS通过防止PARN介导的识别和衰变来维持HIF1a mRNA的稳定性。
为了进一步研究MALR促进的ILF3相分离对肿瘤细胞恶性行为的影响,作者在MALR KO ESCC细胞中过表达了WT-ILF3或DIDR-ILF3(图5H)。作者发现,在缺氧条件下培养的MALR敲除细胞中,WT-ILF3的过表达显著挽救了HIF1a下游分子表达、ILF3点状形成、糖酵解活性、细胞生长和管状形成,但在DIDR-ILF3过表达组中则不是这样(图5H-L)。这些结果表明,MALR-ILF3介导的LLPS促进ESCC的体外恶性进展。
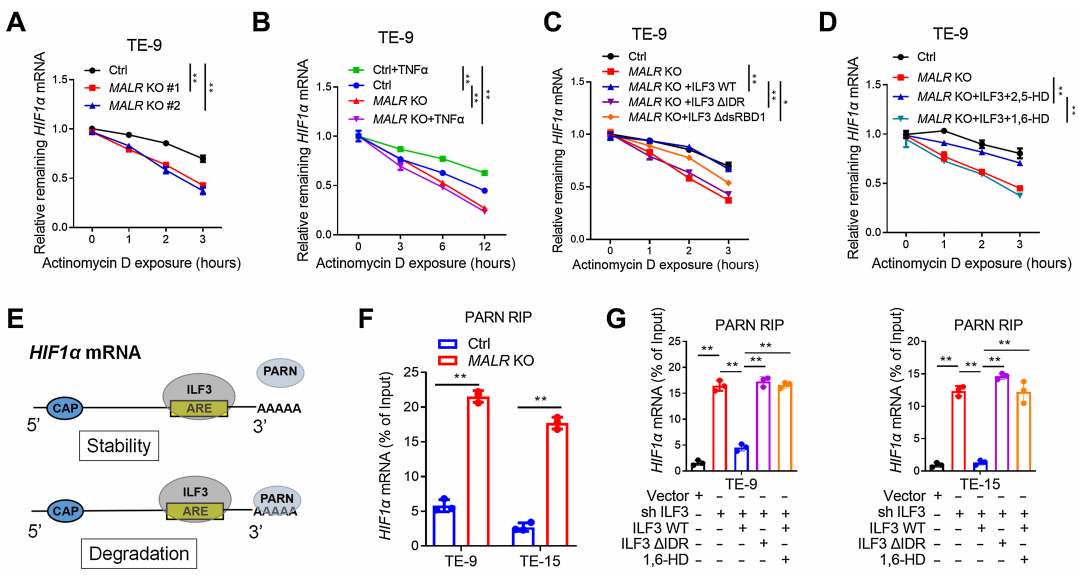
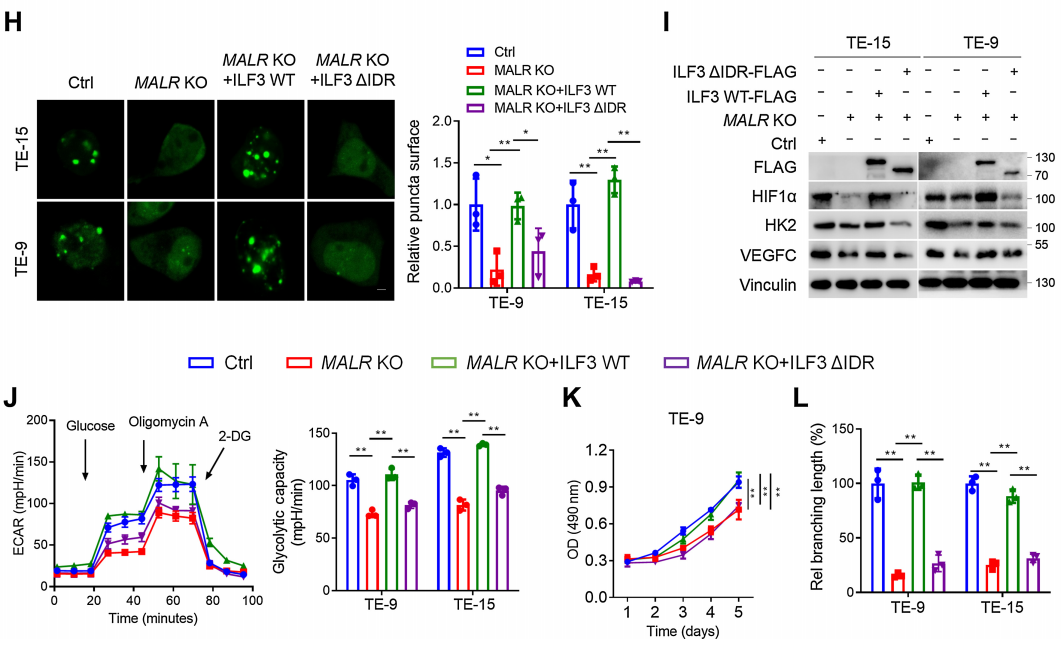
图5. MALR-ilf3介导的LLPS维持了HIF1a mRNA的稳定性
06 MALR-ILF3介导的LLPS促进ESCC的恶性进展
接下来,作者研究了MALR-ILF3-HIF1a轴在体内肿瘤发生中的作用。如图6所示,在ESCC细胞中下调MALR或ILF3显著降低了基于细胞的异种移植肿瘤的生长(图6A和B;补充图S6A和S6B)。此外,Ki-67和CD31(内皮细胞标记)也表明,细胞增殖和血管生成也减少(图6C;补充图S6C)。作者通过IHC和IF染色检测还证实,在异种移植肿瘤组织中,HIF1a表达和ILF3点状形成在MALR或ILF3敲除后显著减少(图6C;补充图S6C)。这些数据与作者在体外的发现一致。作者按照先前报道的方法将ESCC细胞和巨噬细胞注射到NSG小鼠中,增加了异种移植肿瘤的生长、增殖、血管生成和ILF3点状形成,这可以通过在巨噬细胞中消耗TNFa以及在ESCC细胞中消耗MALR或ILF3来消除(图6A-C;补充图S6A-S6C)。值得注意的是,使用MALR抑制剂敲除MALR可显著降低肿瘤生长、血管生成和ILF3点状形成(图6D-H;补充图S6F)。综上所述,这些结果表明MALR在ESCC发展中具有促肿瘤作用。
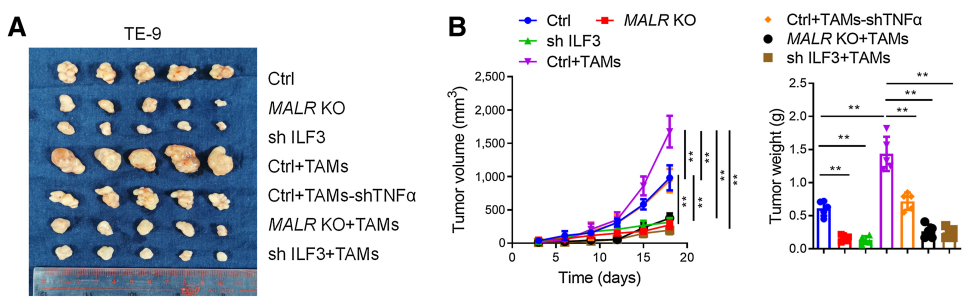

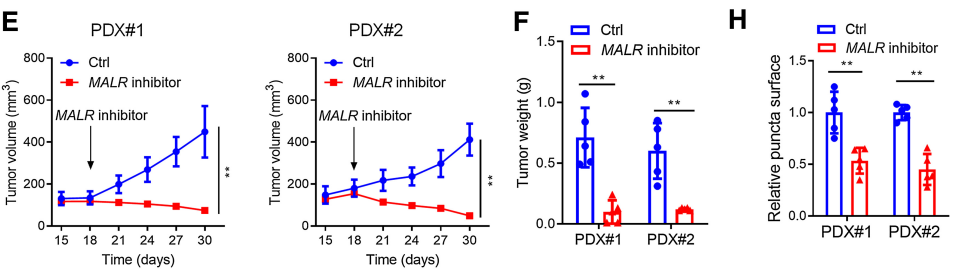
图6. MALR-ilf3介导的LLPS促进体内肿瘤生长
07 ESCC患者中MALR-ILF3-HIF1a轴的临床相关性
为了确定MALR及其上游或下游基因的表达与临床的相关性,作者检查了人类ESCC组织中的它们的表达谱。通过qRT-PCR检测了MALR的表达,并根据中位数表达水平将样本分为MALR-low和MALR-high组。作者进一步通过IHC分析在ESCC组织队列中检查了MALR与TAM标记物(CD68)、肿瘤细胞增殖标记物(Ki-67)和血管生成标记物(CD31)的表达水平之间的相关性(图7A)。MALR-high组表现出较高的CD68、ILF3、HIF1a、Ki-67和CD31表达,而MALR-low组表现出相反的模式(图7B)。值得注意的是,MALR的表达与ESCC组织中ILF3点状的数量呈正相关(图7C)。此外,MALR的表达与HIF1a及其下游基因在ESCC样本中的表达呈正相关(图7D和E)。总之,这些数据表明MALR-ILF3-HIF1a轴在人类癌症的发展中起着重要的作用(图7F)。
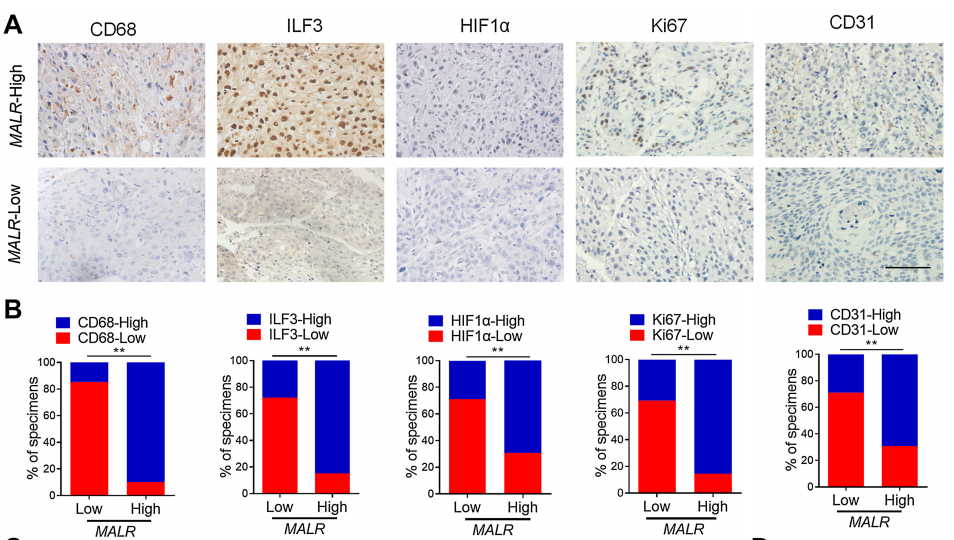
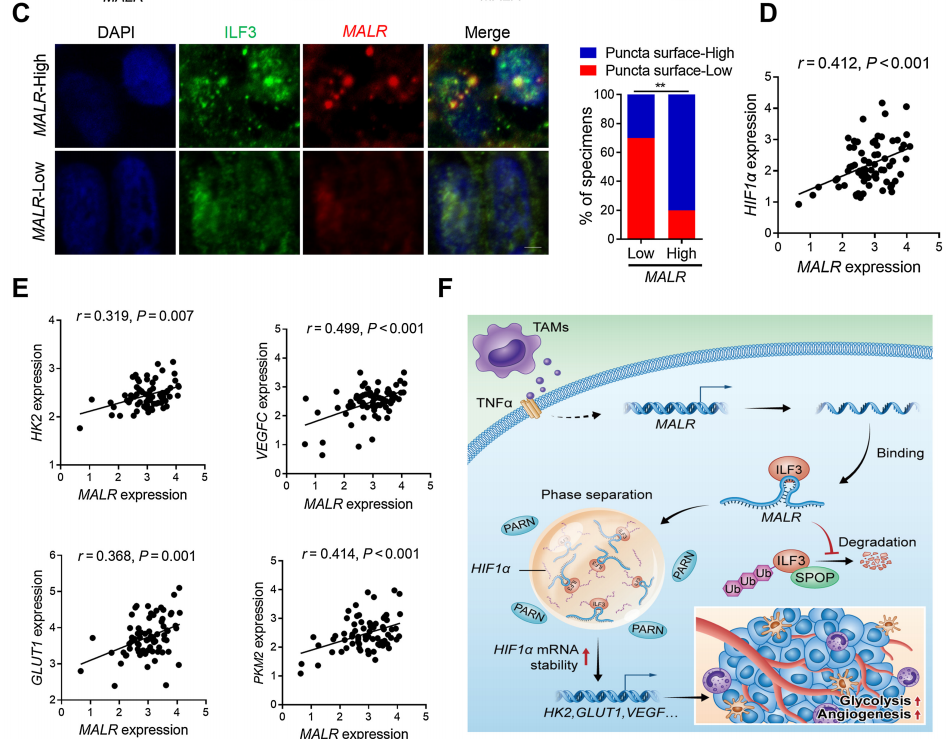
图7. ESCC患者中MALR-ILF3-HIF1a轴的临床相关性
小结
本研究发现lncRNA MALR在食管癌中发挥关键作用。TAMs通过分泌TNFa上调MALR,进而促进糖酵解和血管生成。MALR与ILF3结合增强HIF1a信号通路,促进肿瘤进展。在临床样本中,MALR的高表达与不良预后相关。因此,MALR-ILF3-HIF1a轴可能成为治疗食管癌的潜在靶点。
最新动态
-
07.08
葡萄外泌体“激活细胞清道夫”——SM@G-ELNs协同水凝胶通过MERTK介导的巨噬细胞胞葬作用促进糖尿病伤口愈合(IF=14.3)
-
07.08
三篇顶刊论文,同一个技术选择——BiFC如何破解植物蛋白互作难题?
-
06.24
再登《Cancer Research》!金开瑞双荧光素酶报告系统助力机制研究新突破
-
06.24
登上《Gut》(IF 26.8)!南方医科大学团队揭示葛根类外泌体纳米囊泡靶向肠道细菌治疗类风湿关节炎新机制
-
06.18
亚细胞定位预测与验证结果不一致的原因是什么?
-
06.18
无需抗体、跨物种通用:三篇高水平论文实证DAP-seq如何助力植物转录调控?
-
06.11
打破“单向用药”的局限!IF=11.9《Asian J Pharm Sci》证实:红参外泌体,口服就能双向调控骨代谢
-
06.11
植物外泌体研究前沿:2026年4—5月6项突破性成果系统梳理
-
06.11
IF 26.8!颠覆“越小越好”的认知:南方医科大团队用40微米的柠檬胶囊,让肠屏障主动“开门”抗癌
-
06.11
从零开始研究一个基因,这篇讲透了!
 X
X 






